
The Cost of Acquiring Crossveinless-Ness in Waddington’s Assimilation
Abstract
Neo-Darwinian natural selection theory indicates that sudden, drastic changes in the environment place selective pressure on genetic variants in a population. As time progresses, this pressure sculpts individuals to better fit this new environment. Waddington’s classic experiment was repeated using white-eyed (the w1118strains) flies which produced the crossveinless (cve; disturbed wing crossveins) trait from the parent generation. The F1 generation was split into two selection lines: an Upward Selection Line, that produced more cve in successive generations, and a Downward Selection Line that responded with a consistent but non-linear decline in the percentage of crossveinless. This article will introduce and enlarge observations made on flies with cve; especially the manner in which the Waddington experiment impacts the population. It seems that Waddington evaluated crossveinless just by what it is good for, but not by the price of using it. That is to say, there is an inevitable cost that needs to be paid in order to acquire crossveinless-ness (cve and the associated phenotypes).
Author Contributions
Academic Editor: Ning Shen, Duke University
Checked for plagiarism: Yes
Review by: Single-blind
Copyright © 2017 Ajay Nair, et al.
 This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Competing interests
The authors have declared that no competing interests exist.
Citation:
Introduction
No matter how drastic and sudden, a change in the environment does not bring a change in genetics. Such changes add constraint, by way of increased selective pressure, on those few, rare, genetic variants in the population. It is the repeated nature of selection, with time, that changes the gene frequency in favor of the most suitable individuals. This continues to the point that the whole population comprises more of these fitter variants than the older, less fit wild stock. Waddington’s Genetic Assimilation theory 1 is perhaps an extension to this concept, whereby repeated artificial selection under stress leads to an acquired trait becoming fixed in a population. In 1953, he showed that Drosophila melanogaster (wild-type) flies that were heat-shocked produced a Crossveinless (disrupted posterior crossveins) trait. Through repeated selection of this trait with heat-shock, he not only increased its frequency in the population, but also found that individuals, from the untreated stock, showed the phenotype. However, when it comes to the selection of crossveinless, the repeated selection results in the production of less fit individuals in successive generations. One of the key points about the Natural Selection theory is that nature only favors what is good or beneficial. Crossveinless offers no selective advantage to the heat-shocked flies.
Materials and Methods
Fly Stocks
Two strains of Drosophilamelanogaster were used in the experiment: the wild-type Canton-S flies from Bloomington Drosophila stock center at Indiana University (BDSC) and the local laboratory white-eyed w1118 stock.
Setting Up and Scoring Selection Lines
F1 flies were maintained in sterile disposable culture bottles filled with freshly prepared fly medium until they started pupating (approximately 120 hours AEL). Crossveinless F1 flies were maintained as the Upward Selection Line and the non-crossveinless F1 flies were maintained separately as the Downward Selection Line. Pre-pupal collection was spread over 12-15 time points in a day, for five-seven days. Simultaneously, vials containing prepupae were heat-shocked after 24 hours of incubation (at 25°C) at 40.5°C for 45 min. Following the heat-shock, the vials are left in the incubator (at 25°C) for approximately 5 days, for the flies to eclose (Supplemental data, S1). As they emerge, the flies were scored as either cve or non-crossveinless (non-cve). Unlike the classic experiment 1, 2, special care was taken to ensure the virginity of females before setting up future crosses. Flies (both cve and non-cve) were anesthetized (with CO2), collected, separated (roughly 4-10 hours after eclosion from pupa) into males and females, and stored in yeasted vials until needed. As an added measure, female vials were checked after 3-4 days for any signs of larvae. With this rule for sexing for virgin collection, females were virgin. Once the flies eclosed following heat-shock, they were anesthetized and then scored (S1).
Results
The Waddington Experiment
Following the repetition of the genetic assimilation experiment 1, 2, the upward selection line expressed a near linear increase in the frequency of cve in the population. Whereas the Downward Selection Line displayed a constant decrease in the proportion of crossveinless flies, although the decline was not linear (S1; Figure 1).
Figure 1.Waddington Experiment. (left) graph depicting an overall increase and decrease in the percentage of cve over ten generations in the Upward and the Downward Selection Line. (right)Gradation in the expression of crossveinless. Disturbances in the development of posterior crossveins following appropriate heat-shocking range from being completely missing (as shown in D) to partially disturbed (as shown in A, B, and C; adapted from (2). The bottom two flies show cve phenotype following repetition of Waddington’s experiment in the current study.
The Cost of Acquiring Crossveinless-ness
Although the frequency of the crossveinless allele increased in every generation, there was a cost that crossveinless flies paid in the form of compromised viability. Nothing of the weakened viability was ever mentioned in Waddington’s original work of 1953. But this is not the first time it has been observed. Earlier works 3, 4, 5 have observed compromised viability effects like stockier body and wings, signs of frequent sterility among both sexes, survival of eggs below 50% and dysfunctional male genitalia. Pupal collection in the upward selection line (in the present study) decreased dramatically as one progressed from one generation to the other indicating a severe decrease in the numbers of individuals in the population (Figure 2).
Figure 2.Effect of heat-shock on fly viability. (A) graph showing the drastic decline in the number of pupae collected over ten generations. (B) images showing overall drop in the number of eggs laid and hatched from generation F1 (left) to F2 (right), even after 5 days of incubation. Observations were made from fly-bottles with equal number of flies (100 each).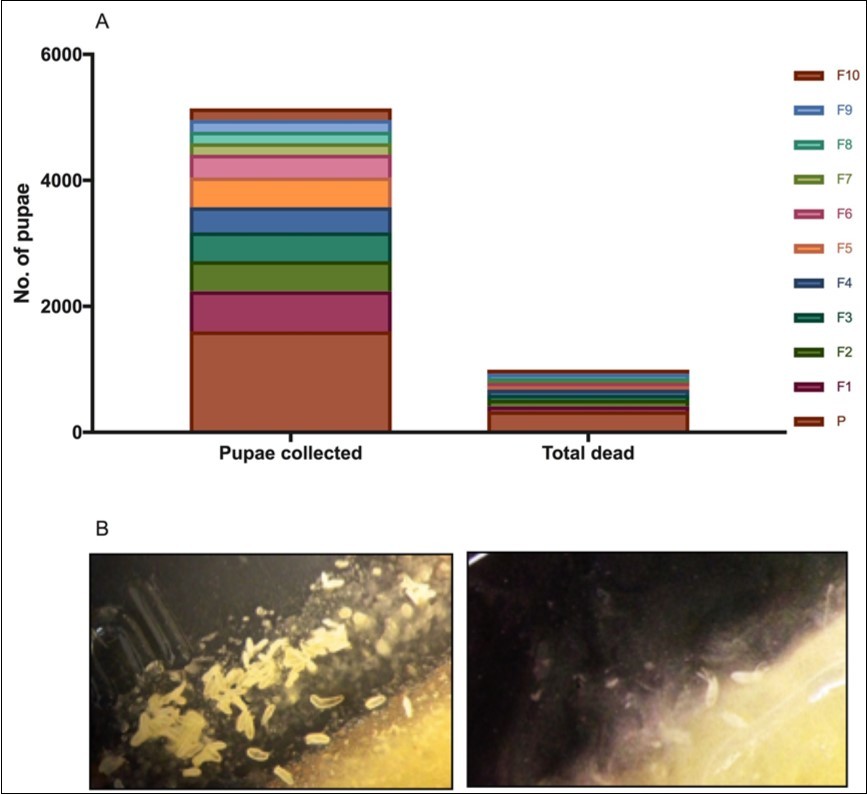
Deviations from Normal Development
During the usual development of Drosophila melanogaster, eggs are laid within 24 hours of turning. Once the eggs hatch, the three instars of larvae (L1, L2 and L3) are formed serially in gaps of ~24 hours. Roughly 120 hours after egg laying (AEL) L3 larvae begin to roam and look for pupation sites (generally as far as possible from food; Figure 3A). Once the top-most part of the fly pot, which is as far as the larvae can pupate, gets occupied, remaining pupariating larvae occupy the next best spot. Eventually, this gives rise to a nearly uniform distribution. Approximately 5 days after pupariation, adult flies eclose or emerge from their pupal cases. However, the heat-shocked population in the upward selection line showed a slight delay in their complete development to adulthood. Firstly, there seemed to be a minor delay in the transformation from instar L2 to L3, and also from instar L3 to prepupae. But one of the most striking and fairly consistent deviations from the typical development was in the stage when L3 larvae roam about in search of appropriate pupation sites. Unlike their normal counterparts (Figure 3B), they pupated either very near to the food or literally on the food (Figure 3C-D). Pupation behavior is important, for it considers the interaction between biotic and abiotic influences 6.
Figure 3.(A) Standard pupation displayed by wild-type flies. Pupation sites, in general, are chosen as far as possible from the food. (B) pupation behavior between cve pupae (left bottle) and non-cve pupae (right bottle). (C) side-view of cve flies pupating quite close to the food and (D) top-view of cve flies pupating on the food. Observations were made from two fly-bottles with equal number of flies (100 each). Equal population size was used to eliminate the effects of population density on pupation behavior.
As mentioned earlier in the section, the flies normally eclose 5 days after the start of pupariation. Except in the case of cve flies, where most of the females eclosed on the fifth day and the males waited a day or two more to emerge out of their pupal cases. Also, in a particular generation females always, and by far, outnumbered males in terms of the percentage of crossveinless (Figure 4). This could mean that cve is either too deleterious to males that many don’t make it to adulthood, or they are more robust towards acquiring the trait. In other words, if the deleterious effects of cve are gender specific (beyond the scope of the current work) then the severity of its effects could explain the lesser number of cve males observed. On the other hand, if the epigenetic landscape for posterior crossvein formation is so robust that it doesn’t decanalize or change upon heatshock, more males would essentially produce normal crossveins rather than the cve phenotype.
Figure 4.cve Female versus cve Male. A column graph showing the mean number of female and male flies in duplicates collected over ten generations in the upward selection line.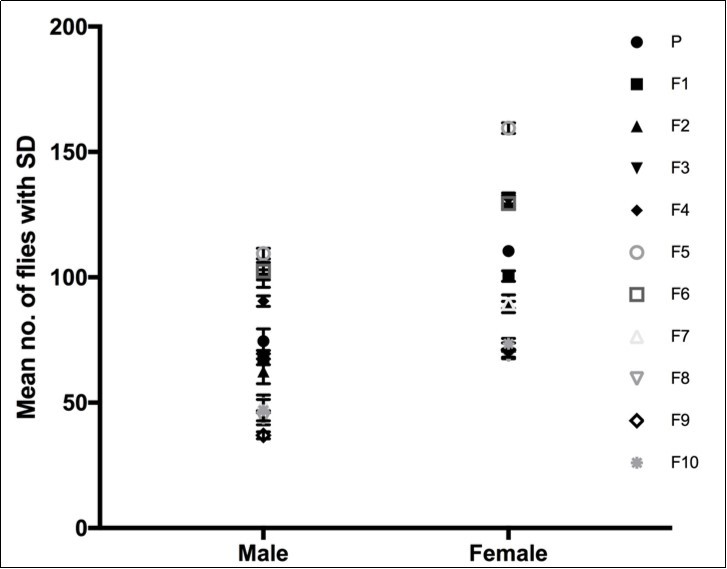
Discussion
The cve Polygene
A reason behind the apparent increase in the frequency of crossveinless, seen in the upward selection line, is that the combinations of genes influencing crossveinlessness are believed to be polygenic in nature 3, 4, 5. That is to say, there is a ‘cve complex’ that influences the fly’s ability to make posterior crossveins, and this complex comprises of a group of common, naturally occurring polygenes distributed randomly over the main chromosomes of Drosophila. Alleles of this complex act additively in lowering the ability to make posterior crossveins in selection lines. Repeatedly selecting cve individuals in successive generations improves the probability of having these random alleles, and this eventually increases the percentage of cve. Also, the alleles of this polygenic system have no visible effects by themselves, but do so when they act in combination. Any of these several combinations may produce the crossveinless response. This is why there is always a range in the expression of the character, instead of just one degree of severity all the time. Although every cve response from a heat-shocked population could look like a simple disturbance in the posterior crossvein, it is more complex. A disturbed posterior crossvein could result from either incomplete expression of crossvein promoting signals or the improper inhibition of intervein region promoting signals at the vein/intervein boundary. So in each cve fly, if one assumes there is a 50% chance of producing posterior crossvein defects from either of the above-said sources. Within each of the 50% probability, a crossvein (as mentioned earlier), might be either disturbed with a small nick at the point where it meets the longitudinal veins, three-fourth missing, or completely absent in the wings (each having a ~25% chance of occurrence). Any of these three differences have an equal chance of appearing in the population. Furthermore, any of the above-mentioned three defects could be in either one wing or in both wings (so that is like a 12.5% probability). Although it has been observed that females show more susceptibility to the response, the percentage of males showing cve is not negligible. The net probability of crossveinless-ness in a population, therefore, comes to approximately 6.25%; that is to say each grade of cve only has a 6.25% (rough estimate) chance of occurring again in the same generation. So, this is the extent to which the trait varies in the population, and in doing so, it further validates the fact that crossveinless is affected by a combination of many random factors spread out in the genome (Figure 5).
Figure 5.A simplified flowchart that illustrates the gradation seen in the expression of crossveinless in the Waddington experiment.
Viability Compromised for Optimum Homeostasis.
We know that energy is used for two vital requirements in a living system: one for maintenance and the other for reproduction 7. Under usual circumstances, the balance between the two is upheld for the perpetuation of life, but why? During favorable times, energy is abundant in the system. And this leads to increase in the net Entropy. To circumvent this, energy is equally distributed, thereby minimizing the entropy and achieving homeostasis. On the other hand, during unfavorable times like a recurrent heat-shock, the usable energy (Gibbs free energy, G) depletes, leading to increase in disorderness and decrease in homeostasis. To tackle this situation, the system might well take an approach that may seem detrimental initially but it will help achieve an overall homeostasis, or one that allows survival, but has viability/fitness consequences later. The system tilts the balance in favor of maintenance, and thus reproduction is compromised (until favorable conditions return). But an extreme measure such as this would ensure that the development of the progeny doesn’t suffer during in optimum conditions, due to the resultant energy insufficiency. In doing so, homeostasis is attained for the time being. In the present study, this may explain why there is a reduction in the number of eggs that hatch, to consistent delays in the development of the larval and pupal stages, to pupation sites chosen not far away from the food source, to lesser number of adults being fertile. To attain homeostasis and ensure interim survival, the system perhaps compromises on fitness. So, considering the impacts heat-shock has on the fitness and the development of flies, one realizes that crossveinless is much more than the mere disruption of crossveins.
Conclusions
Repetition of the Waddington experiment showed that just like the original experiment, the fly population strongly to the recurrent heat-shock and selection. Although the classic experiment made the selection of crossveinless look something as straightforward as merely heat-shocking pupae at specific conditions, it turns out there is a cost associated with acquiring crossveinless. It is understood that crossveinless as a phenotype does not offer any benefit to the fly 1, but it surely seemed to be deleterious (in the present study) to the biological fitness of flies
Supplementary data:
Acknowledgements
We would like to thank GRAVIDA, New Zealand and the University of Otago, New Zealand for providing funding and research facilities that was key to the successful completion of this work.
References
- 2.Nair A, P K Dearden. (2016) A Systems View of Waddington’s Genetic. , Assimilation.International Journal of Bioinformatics and Biological Systems 1(1), 10-17.
- 3.R D Milkman. (1959) The genetic basis of natural variation. I. Selection for Crossveinless polygenes in new wild strains of Drosophila melanogaster.Genetics(50):. 625-632.
- 4.R D Milkman. (1962) Temperature effects on day old Drosophila pupa.The. , Journal of General Physiology(45): 777.
- 5.R D Milkman. (1960) The genetic basis of natural variation. II. CrossveinsinDrosophilamelanogaster.Genetics(45): 3548